筋肉の解剖学とVBT

筋肉の解剖学と筋力向上の理解は密接に関連しています。この記事では、解剖学的観点から筋肉の収縮メカニズムを解説し、速度を重視したトレーニングの原理と目的が、それといかに直接結びついているかを説明します。
VBTと筋収縮
以下で述べる内容はすべて、筋力トレーニングや、特に速度を重視したトレーニングとの関連性を理解するために、筋の解剖学と生理学について非常に具体的な知識を提供することを目的としています。以前の投稿では、力-速度プロファイルについて触れました。力-速度関係とは、単に、筋肉の長さが変化する速度(外部負荷や他の筋肉によって制御される)と、その筋肉が発生する力の大きさとの関係を指します。 個人の筋組織の特性によって、力-速度プロファイルの曲線がどのような形になるかが決まります。また、その曲線は、収縮ごとに動単位をより多く動員することや、収縮ごとの発火頻度を高めることによって、さらに変化させることができます。そして、これら2つの変数は、トレーニングによって、特に速度ベースのトレーニングのように、目的を持って特化したトレーニングを行うことで変化させることができるのです。
Perch(筋力測定)機器による即時的かつ客観的なフィードバックにより、リフトの意図が数値化されるだけでなく、経時的な変化も追跡されます。これらのデータポイントにより、コーチは選手の筋肉の奥深くで実際に何が起きているのか、その一端を捉えたり、基礎的な理解を得たりすることができます。努力の度合いに数値を割り当てることで、選手はさまざまな負荷レベルにおける筋収縮の感覚を理解しやすくなり、自身の身体との調和を深めるよう促されます。筋肉により大きな力を生み出せるように鍛えることは、単純ではありますが、決して容易なことではありません。 プログラムでは、アスリートに以下のことを指導する必要があります:
- 収縮ごとに、より多くの運動単位を動員する
- すでに活動している運動単位群の発火頻度を高める
速度を重視したトレーニング技術が、筋力トレーニングやスポーツパフォーマンス向上のさまざまな場面で一般的になるにつれ、目標達成までのスピードが加速し、アスリートは自身の潜在能力を最大限に引き出すことができるようになります。以下では、筋肉が収縮する理由と仕組みについて解説します。
筋収縮の種類
筋肉の収縮には4つのタイプがあります:
- 等尺性収縮:筋肉の長さを変えずに張力を生み出す
- 等張性収縮:筋肉の長さが変化しても、一定の張力を生み出す。
- 同心収縮:筋肉の張力が、それに抗する外力を上回り、収縮に伴い筋肉が短縮する
- 伸張性収縮:筋の張力は、それに抗する外力よりも大きくなく、収縮中に筋が伸長する。
骨格筋の解剖学
(反射を除く)すべての骨格筋の収縮は、脳から始まります。電気化学的な信号が神経系を通じて、複数の筋線維を支配する運動ニューロンへと送られます。個々の筋肉の実際の解剖学的構造は、以下の通りです:
![科学者シンディ [6] の解説を基に、骨格筋の構造を層ごとに詳しく見ていきます。](https://www.catapult.com/wp-content/uploads/2026/04/664f34c5468e7068a06f718e_63ce4a75736e68eeaec04190_fiber.jpg)
小さいものから大きいものへと順に、筋肉組織の層は以下の通りです:
筋原線維:筋肉の収縮を司る、最も小さく、基本的かつ機能的な単位。互いに絡み合った繊維(アクチンとミオシン)から構成され、筋線維の横紋を形成している。1つの筋原線維の中には、多数の筋原線維が存在する。
筋原線維:太い筋原線維と細い筋原線維(アクチンやミオシンと呼ばれる収縮性タンパク質、およびトロポニンやトロポミオシンと呼ばれる調節性タンパク質)から構成される、筋線維内の細長く平行な単位。筋小胞体(SR)に囲まれている。
筋線維:多数の筋原線維を含む、長い円筒形の細胞。筋膜に囲まれている。筋細胞とも呼ばれる。
筋膜:各筋線維を包む細胞膜または原形質膜。
内筋膜:単一の筋線維を包む、最も小さな結合組織。
筋束:筋膜に囲まれた筋線維の束。
ペリミシウム:筋束構造の中で、複数の筋線維を包み込む中程度の大きさの結合組織。
筋膜:結合組織の中で最も大きなもので、弾力性のある繊維質の鞘であり、筋肉全体を包み込むことで、筋肉の構造を維持しつつ、周辺の他の組織や臓器とは独立して動くことを可能にしている。
筋膜:筋肉全体を覆い、筋膜下組織の層の上に位置する、厚い結合組織の層。
神経筋接合部
神経筋接合部(筋神経接合部や運動終板とも呼ばれる)は、本質的に運動ニューロンと筋線維の接点間に形成される化学シナプスである。その最も基本的な単位は運動単位と呼ばれ、単一のα運動ニューロンと、それが支配するすべての筋線維から構成されている。その構造は以下の通りである:
![運動単位の図解。Gardiner [2] より引用・改変。](https://www.catapult.com/wp-content/uploads/2026/04/664f34c5468e7068a06f7196_63ce4a75736e68eaa3c04191_motor-unit.png)
運動ニューロンは、細胞体、樹状突起、細胞核、軸索(ミエリン鞘で覆われている)、および軸索終末から構成されています。 軸索はシナプス小球またはボタン(シナプス前側)で終わり、そこがシナプス接合部を形成する場所であり、ボタンの末端と標的細胞(シナプス後側)の始点との間にはシナプス間隙が存在する。 骨格筋において、シナプス後側の標的細胞には、受容体で覆われた一連のシナプス皺があります。以下に、神経筋接合部で起こる現象を段階的に要約します:
- 活動電位が運動ニューロンを伝わり、シナプスボタンがアセチルコリンと呼ばれる神経伝達物質をシナプス間隙に放出する。
- アセチルコリンは、シナプス後側の接合部ひだにあるアセチルコリン受容体に結合する。
- 結合すると、イオンチャネルが開き、陽イオンのナトリウム(Na)がシナプス後細胞内に流入する。これにより細胞は脱分極し、終板電位が生じる。
- 脱分極により電圧依存性ナトリウム(Na)チャネルが開き、終板電位が活動電位へと変化する。
- 活動電位は筋線維に沿って伝わり、興奮収縮連関を通じて筋線維の収縮を引き起こす。
![Gonzalez-Friereら[3]による「神経筋接合部の構造」](https://www.catapult.com/wp-content/uploads/2026/04/664f34c5468e7068a06f7199_63ce4a75da2bff4eee00089a_junction-arch.jpg)
興奮収縮連関
興奮収縮連関とは、シナプス後側で起こる一連の事象であり、その概要を以下に段階的に示します:
- 終板電位の脱分極によって引き起こされた活動電位は、細胞膜の残りの部分を伝わり、細胞表面を横切って広がる
- 活動電位は、筋小胞体(SR)に接しているT管と呼ばれる構造へと伝わる
- 活動電位により、粗面小胞体(SR)の末端システィーナから細胞質へとカルシウム(Ca)が放出される
- その後、カルシウムイオンはトロポニンと結合し、トロポミオシンを移動させて、アクチン上のミオシン結合部位を露出させる。
- ミオシンヘッドはアクチンと架橋を形成し、筋収縮を開始する
- ATPはミオシンヘッドに結合し、ミオシンヘッドを解放してリセットさせる
- 酵素反応によってCaがSRに戻されると、弛緩が起こる
![神経筋接合部で始まる興奮収縮連関の概要。Scientist Cindy [6] を改変](https://www.catapult.com/wp-content/uploads/2026/04/664f34c5468e7068a06f7191_63ce4a75b086d4214f082232_excite-contract.png)
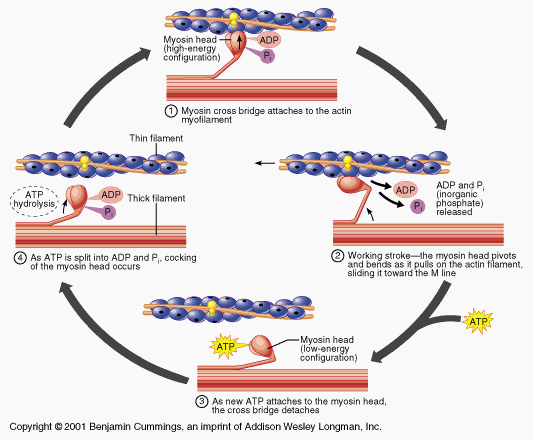
滑動フィラメント説
「滑走フィラメント説」とは、筋肉収縮の最も基本的なメカニズムを指すものです。興奮収縮連関との共通点も一部ありますが、ここではその概要を順を追って説明します:
活動電位は、筋細胞内へのカルシウムの放出を促進する
Caは(それまでアクチンに結合していた)トロポニンに結合し、それによってアクチンからトロポミオシン鎖が除去され、ミオシンが結合できる部位が確保される。
ミオシンの球状頭部が、ADP + Pの形態をとったATPを用いて利用可能なアクチン結合部位に結合すると、「パワーストローク」が発生し、アクチンフィラメントが中心部、すなわちMラインに向かって引き寄せられる
その後、新しいATPがミオシンに結合し、それによって形成されたクロスブリッジがアクチン結合部位から離脱する。
ATPがさらに存在し、新たなクロスブリッジを形成できる場合は、筋繊維は収縮を継続できるが、そうでない場合は弛緩し、CaはSRへと戻される。
骨格筋の収縮における違い
筋収縮は(前述の通り)活動電位によって制御されており、一般的には次のように分類される:
- ツィッチ:筋線維そのものの中で生じる単一の収縮・弛緩サイクル
- (波状収縮):連続する複数の微小な収縮が重なり合い、より大きく強力な筋収縮が生じる現象
- 破水:複数の収縮が重なり合って、持続的で強い収縮を引き起こすもので、破水している場合も、破水していない場合もある。
最も基本的なレベルでは、骨格筋で発生する力の大きさを変える方法は2つしかないということを覚えておくことが重要です:
- 収縮ごとに、より多くの運動単位を動員する
- すでに活動している運動単位群の発火頻度を高める
動員可能なすべての運動単位が動員され、最大頻度で収縮している状態が、1回最大挙上重量(1RM)となります。身体は負荷がかかると、常に現在使用中の運動単位を消耗させるよりも、より多くの運動単位を動員することを選びます。収縮の持続時間や程度も、以下の方法による運動単位の動員によって調節されます:
- 活動中の運動ニューロンの数を増やす
- まず最も小さく弱い運動単位を活性化させ、その後、より大きな運動単位を活性化させる
結論
Perch、あらゆる事柄の「なぜ」を理解することを強く推奨しています。そのため、筋肉を的確に鍛え、総合的な運動能力を高めるためには、速度を重視したトレーニングがあらゆるウェイトトレーニング環境において不可欠であると考えていますが、これを真に理解するためには、筋肉の解剖学を学ぶことが重要だと考えています。この記事が皆様のお役に立てれば幸いです!
その他の関連記事!
VBTの基本についてもっと知りたいですか?Perch用語集をチェックしてみてください!
VBTで筋肉がどのように成長するのか気になりますか?筋肉の成長とベロシティ・ベースド・トレーニングに関する記事をご覧ください!
フォローしてください!
速度を重視したトレーニングに関するコンテンツやヒント、コツ、ツールなどを随時更新していきますので、ぜひチェックしてください。また、Twitter、Instagram、LinkedInでのフォローや、Facebookでの「いいね!」もお忘れなく。
出典
- Baechle, T., Earle, R., & 全米ストレングス&コンディショニング協会(米国) (2008). 『ストレングストレーニングとコンディショニングの基礎』(第3版). イリノイ州シャンペーン:ヒューマン・キネティクス.
- ガーディナー, P. (2011). 『神経筋運動生理学の応用』(『運動生理学の応用』シリーズ)。イリノイ州シャンペーン:ヒューマン・キネティクス。
- Gonzalez-Friere, M., Rafael, de C., Stephanie, S., & Luigi, F. (2014年8月). 神経筋接合部の構造。 2019年10月23日にhttps://www.researchgate.net/figure/The-architecture-of-a-neuromuscular-junction-NMJ-A-B-The-NMJ-is-composed-of-three_fig1_265056822 から取得。
- Gray, H., Williams, P., & Bannister, L. (1995). 『グレイの解剖学:医学・外科学の解剖学的基礎』(第38版)。ニューヨーク:チャーチル・リビングストン。
- Scanlon, V. & Sanders, T. (1999). 『解剖学・生理学の基礎』(第3版). フィラデルフィア:F.A. Davis.
- Scientist, C. (発行年不明). 「筋肉と反射の実験」。2019年10月23日にhttps://www.scientistcindy.com/muscles-and-reflexes-lab.html から取得。